分子机制研究是大家最为头痛的主题,今天我们就来分享一下研究分子机制的经验和套路:
(1)信号通路;
(2)明星分子;
(3)中心法则;
(4)工具分子;
(5)具体位点;
(1)信号通路
信号通路千千万,找到一条是关键,通路关系太复杂,讲不清楚就完蛋!
确定信号通路的方法有很多,特别是当我们已经把一个基因的细胞和动物功能阐述清楚后:
1)通过文献报道或者已知经验,比如分子A的功能是影响细胞自噬,那么像PI3K-Akt-mTOR通路等就是我们可以直接通过阅读综述获取的;
2)通过RNA-seq或者芯片或者其它组学手段,筛选差异基因并对差异基因进行pathway或者GSEA富集分析,从而找到可能被基因A调控的信号通路;
3)通过数据库预测,比如基于与基因A存在共表达关系来分析周围基因所参与的通路,从而得到A可能参与的通路,我们可以通过cbioportal/GEPIA对TCGA收录肿瘤中的信息进行分析,也可以分析GEO等非肿瘤的共表达结果,甚至有一些网站直接收录了这种共表达关系。
(2)明星分子
明星分子选得好,文章分数才能高,中游节点最常选,上下连接有得搞!
明星分子的概念大而且含糊,尽管通路是一个大的网络,但是总体上上下游关系还是存在的,如果简单来划分可以分为上游,中游和下游三段,三者之间没有严格的界限,一般的通路常常出现分子类型有:配体与受体,激酶/磷酸酶(去/泛素化酶)与底物,转录因子和靶基因等,以TGFβ-Smad通路为例,如果把细胞膜和核膜作为上中下分类的区分点,可以粗略的这么划分和标注:
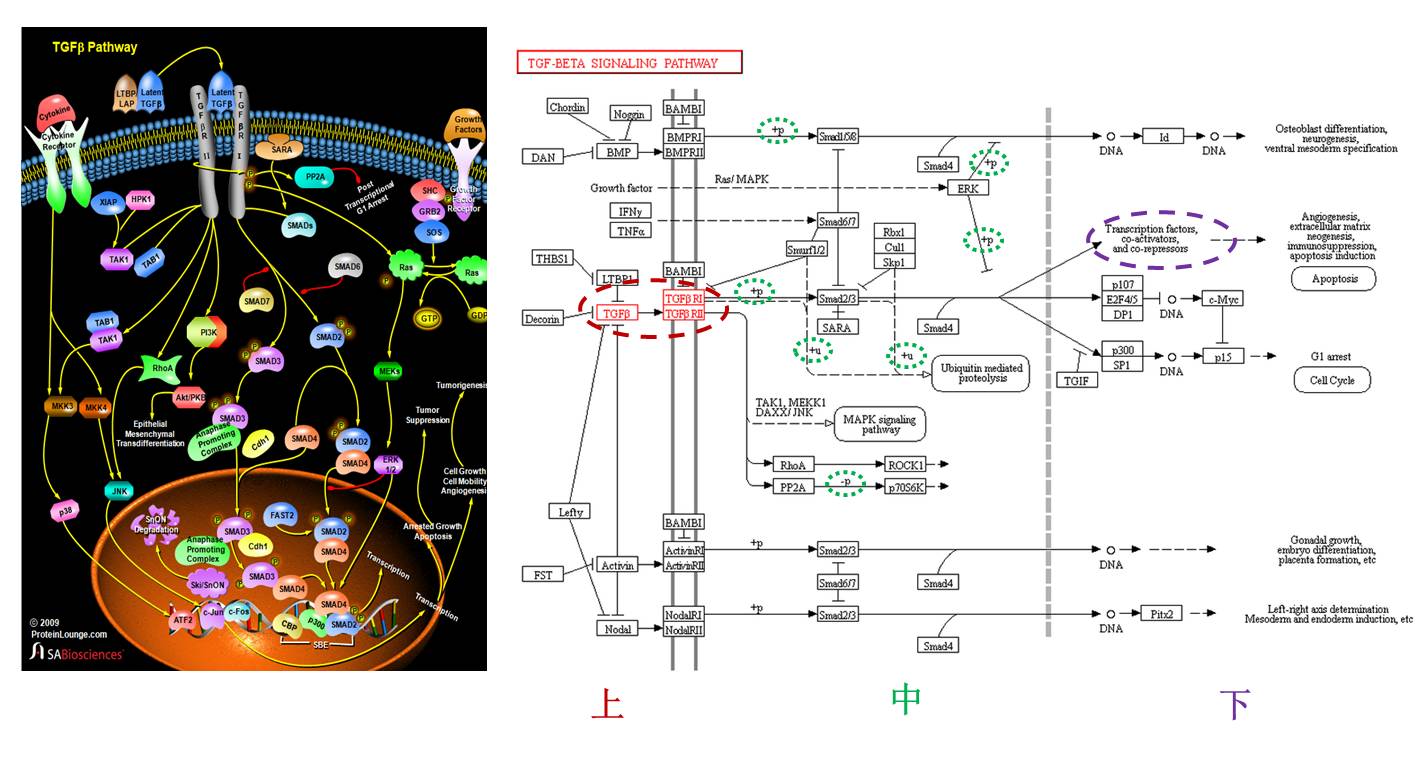
那么在这个通路里面什么分子是明星分子呢?一般来说,大家选的重点集中在中游这个范围内,下游的分子(比如图中的p15)一般用来作为细胞表型(细胞周期)的证据,可能考虑到膜蛋白研究的复杂和困难性,大家也很少选择上游的分子,所以中游的分子就成了明星分子选择的重点了,特别是通路的名字叫做TGFβ-Smad,所以很多研究就是围绕Smad这个蛋白展开,比如Smad这个家族的蛋白即是转录因子,可以通过结合靶基因的DNA调控分子表达,其本身又可以被磷酸化发挥功能,所以又是激酶-底物这一关系中的底物这一角色,因此激酶-Smad-磷酸化的Smad(转录因子)-靶基因就成了一个信号轴。
(3)中心法则
中心法则像楼梯,三个水平N多级,从大到小逐层找,推理验证少不了!
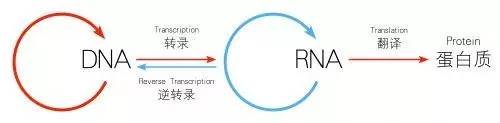
当我们选好明星分子以后,调控层次就是要考虑的问题了,这里说的调控层次就是围绕中心法则展开对明星分子本身的探索,我们知道中心法则涉及到DNA-RNA-蛋白三个层面,根据调控的层次一般我们又分为转录水平和转录后水平,转录我们知道是发生在细胞核里面的。所以我们看到某个分子定位在细胞核时我们一般就从转录水平进行考虑,比如转录因子对DNA序列的结合,DNA甲基化、组蛋白修饰等等,而当RNA被转录出来后要经过加工过程,比如可变剪接,miRNA介导的降解,mRNA翻译成蛋白,蛋白合成后会被磷酸化修饰或者泛素化修饰从而激活或者降解等等,这些是转录后的水平。那么我们在确定明星分子后,一般来说需要首先确定的是变化发生在哪个层面,是RNA还是蛋白,是蛋白的产生(翻译)、降解(比如泛素化)还是活化(比如磷酸化),从而把研究的焦点集中到明星分子的某个过程,这个层面是围绕一个明星分子的前世今生来展开的。
(4)工具分子
一个结果多个因,工具分子一大群,具体哪个有作用,IP验证指乾坤!
确定了明星分子在哪个层面被调控后,就要找相应的工具分子了,工具分子是指像酶(比如激酶、泛素酶、甲基化酶)、转录因子、翻译因子、剪接因子等为主的一系列蛋白,每个系列的工具分子一般都有很多,比如泛素酶就有成百上千个,那具体是哪一个导致明星分子被泛素化呢?以免疫沉淀(IP)技术为基础延伸出来的各种分子就是用来找这类工具分子的最常用策略,比如通过IP加质谱鉴定与明星分子作用的泛素化酶,通过RNA pulldown加质谱鉴定与RNA结合的工具蛋白(如翻译因子和剪接因子),通过RIP-seq来寻找与蛋白结合的RNA,通过ChIP-Seq来寻找与蛋白结合的DNA序列(如靶基因)等等。找到工具分子后,作用的信号轴就有了:某基因(因素)通过(4)工具分子从(2)明星分子(3)某个层面调控(1)某条信号通路。
(5) 具体位点
分子整体有作用,具体位点讲不清,拆开组合来验证,整个世界都安静。
这一步就是要进一步把明星分子和工具分子进行拆解,工具分子揍了明星分子一顿,具体是揍了一拳还是踢了一脚,工具分子的一拳打在了明星分子的脸上还是肚子上,这个是两个要交代清楚的问题,所以就要把工具分子和明星分子中发挥功能的位点、序列或者结构域找到,找到后就可以更加清楚的描述了:工具分子左拳打在了明星分子头上,具体位置是打在了太阳穴上,把明星分子打死了,这样导致一个以明星分子为中心的网络(信号通路)出现崩溃或者瘫痪,异常(疾病)就出现了。所以我们在文章中常常看到要把蛋白根据结构域进行分段,把RNA根据序列长度进行分段,然后再看他俩之间还有没有作用。比如大家当做笑话来看的一个实验,一研究人员把一只蜘蛛放在桌上,对它大喊一声,蜘蛛吓跑了;把蜘蛛的腿全撕了,对它大喊一声,蜘蛛没跑,所以结论是:蜘蛛的耳朵是长在腿上的。
看看人人网的大神是怎么设计的:
Fig 1. 蜘蛛形态学的描述,重点引用大量的文献提示蜘蛛的听觉系统可能在腿上。
Fig 2. 蜘蛛对声音的反应性实验。
a. 对蜘蛛大吼---->蜘蛛跑了
b. 把蜘蛛的腿用布包起来---->蜘蛛不动
c. 把蜘蛛的脚一条条切掉,再对蜘蛛大吼----->蜘蛛移动距离随着切掉的条数增多而降低!
d. 把切掉的蜘蛛腿装回去!!再对蜘蛛大吼--->蜘蛛恢复了部分的反应性,p<0.0000000001
Fig 3. 昆虫的耳朵都保守地长腿上。
a. 用小强重复Fig 2的实验;
b. 用蝴蝶重复Fig 2的实验
c. 继续用其他目的昆虫重复实验
---------------------------------------
做到这里,你可以发CNS的letter了,要发到article的级别,继续吧!
---------------------------------------
Fig 4. 大范围的调查实验,腿越多的动物对声音的反应性越强啊,关联性分析啊。
Fig 5. 基因组测序+分析啊,发现一大堆跟长腿有关的基因啊~(这里可以顺便发一篇nature biotechnology或者nature genetics神马的),然后做基因敲除,敲了不长腿的蜘蛛,吼了不反应啊~!插入更多长腿的基因,长出16条甚至32条腿,吼了之后跑更远了亲!!
Fig 6. 你做出来就拿诺奖了!把这些基因用于先天性失聪病人的治疗,尼玛又听见了啊!!(这里又顺便发一篇New England或者JAMA神马的),副作用是可能长一点点蜘蛛腿…………