郑重声明:原文参见标题,如有侵权,请联系作者,将会撤销发布!


Cell, (2015): 456-492
SUMMARY
我们提出了幼年大鼠体感皮层微电路的初稿数字重建。重建使用细胞和突触组织原理从稀疏的实验数据中通过算法重建详细的解剖学和生理学。客观的解剖方法定义了包含~31000个神经元的0.29 ± 0.01 mm3的新皮质体积,膜片钳研究确定了55个特定层的形态学和207个形态电神经元亚型。当数字重建的神经元被定位在体积中并且突触形成仅限于生物布顿密度和每个连接的突触数量时,它们的重叠轴形成~800万个连接和~3700万个突触。模拟在没有参数调整的情况下再现了一系列体外和体内实验。此外,我们发现了一系列网络状态,这些状态具有从同步到异步活动的急剧转变,受生理机制的调节。围绕这一转变动态重新配置的网络状态谱支持各种信息处理策略。
INTRODUCTION
自从Santiago Ramón y Cajal对新皮质的开创性工作(DeFelipe and Jones, 1988; Ramón y Cajal, 1909, 1911)以来,大量研究试图揭示其多层次的解剖组织(神经元的类型、突触连接、分层、新皮质区域内和之间的传入和传出投射等)和功能特性(神经元反应特征、突触反应和可塑性、感受野、功能性新皮质柱、紧急活动图、新皮质区域之间的相互作用等)。然而,我们的知识仍然存在很大差距,特别是在细胞和突触水平上新皮质的解剖和生理组织。
具体而言,虽然神经元已根据其电生理行为进行分类(Connors and Gutnick, 1990; Kasper et al., 1994; McCormick et al., 1985),但不同钙结合蛋白和神经肽的表达(Celio, 1986; DeFelipe, 1993; Gonchar and Burkhalter, 1997; Kawaguchi and Kubota, 1997; Toledo-Rodriguez et al., 2005)和形态特征(Kisvárday et al., 1985; Larkman, 1991a; Tamás et al., 1998; Wang et al., 2002),关于神经元类型的客观和全面分类仍未达成共识。尽管不同神经元的蛋白质和遗传标记分布(Grange et al., 2014; Hendry et al., 1989; Kawaguchi and Kubota, 1997; Meyer et al., 2002; Toledo-Rodriguez et al., 2004)和已经描述了一些形态学和电学分类神经元的相对比例(Beaulieu and Colonnier, 1983; Cauli et al., 1997; Hendry et al., 1984; Meyer et al., 2010a; Rudy et al., 2011),但我们缺乏全面了解每层中每种类型的神经元的数量。自从配对记录技术出现以来,一些研究已经描述了某些类型神经元之间突触连接的解剖学和生理学特性(Cobb et al., 1997; Feldmeyer et al., 1999; Frick et al., 2008; Gupta et al., 2000; Mason et al., 1991; Reyes et al., 1998; Thomson et al., 1993),但很大一部分还有待研究。尽管使用逆行和顺行示踪剂和跨突触病毒载体进行标记,但使用阵列断层扫描成像和使用电子显微镜进行饱和重建使得开始绘制单个新皮质神经元的突触前和突触后神经元成为可能(Boyd and Matsubara, 1991; Callaway, 2008; Glenn et al., 1982; Kasthuri et al., 2015; Killackey et al., 1983; Micheva and Smith, 2007; Micheva et al., 2010; Wickersham et al., 2007),我们既不知道与任何特定神经元类型相关的突触前和突触后神经元的数量与类型,也不知道它们与其直接邻居形成的突触的数量和位置。
在功能层面上,已经对新皮质切片中的紧急行为进行了许多研究(Cunningham et al., 2004; Mao et al., 2001; McCormick et al., 2003; Sanchez-Vives and McCormick, 2000; Yuste et al., 1997)、相关活动(Hasenstaub et al., 2005; Livingstone, 1996; Salinas and Sejnowski, 2001; Shu et al., 2003; Silberberg et al., 2004; Singer, 1993),以及单个神经元跨皮质层的功能影响(Sakata and Harris, 2009; Schroeder and Foxe, 2002; Silva et al., 1991; Steriade et al., 1993)以及体感和其他皮质区域的体内活动(Chen et al., 2015; Klausberger et al., 2003; Leinekugel et al., 2002; Luczak et al., 2007; Reyes-Puerta et al., 2015; Wilson et al., 2012),然而,我们仍然缺乏对细胞和突触机制和不同层在最简单的行为中的作用,例如相关和不相关的单神经元活动,更一般地说,同步和异步群体活动。例如,众所周知,不同类型的神经元通过具有不同动力学和强度的突触连接,战略性地位于神经元树突、胞体和轴突的不同位置,但这种组织的功能意义仍不清楚。抽象出这一级别的生物学细节的计算方法无法解释这种复杂的细胞和突触组织的功能意义。尽管未来的实验研究无疑会增进我们的知识,但仅凭实验映射是否可以提供足够的数据来回答这些问题,仍存在争议。
在此,我们提出了一种互补的算法方法,该方法使用可用的稀疏数据在所有层上重建神经元微电路,并利用生物学原理和数据集之间的相互依赖性来预测丢失的生物学数据。作为测试案例,我们以数字方式重建了2周大的Wistar (Han)大鼠后肢体感皮层第1层至第6层的少量组织。选择该模型系统不仅是因为它是新皮质中最全面的特征之一,还因为其细胞和突触组织的实验数据很容易获得,并且验证实验相对容易执行。简而言之,我们从体外脑切片中记录和数字重建神经元,并根据成熟的形态类型(m型;图1A)对神经元进行分类,根据实验将神经元定位在客观定义尺寸的数字体积中。估计它们的特定层密度(图1B),并重建了神经元之间的连接(图1C)。然后使用Petilla公约(Ascoli et al., 2008)中提出的分类的扩展版本将神经元分为电类型(e型),并生成了捕获每种类型的特征电行为的模型。(图1D);同样,对突触进行建模以捕获特定突触类型(S型;图1E)的特征突触动态和动力学。最后,我们使用实验数据构建了一个虚拟切片并重建了丘脑输入(图1F;Meyer et al., 2010b)。
这种方法产生了微电路的初稿数字重建,并针对重建中未使用的大量实验数据集进行了验证。结果表明,无需测量所有可能的生物参数即可获得神经微电路的密集图,并指向所需的最少数据集,即战略数据。整合互补但稀疏的数据集也使得调和文献中的差异成为可能,至少部分解决了数据质量和可重复性的问题。探索重建微电路的一些紧急行为的模拟重现了许多先前的体外和体内发现,并为新皮质微电路的设计和功能提供了见解。实验数据、数字重建和模拟结果可在新皮质微电路协作门户(NMC门户;https://bbp.epfl.ch/nmc-portal;参见Ramaswamy et al., 2015)上获得。
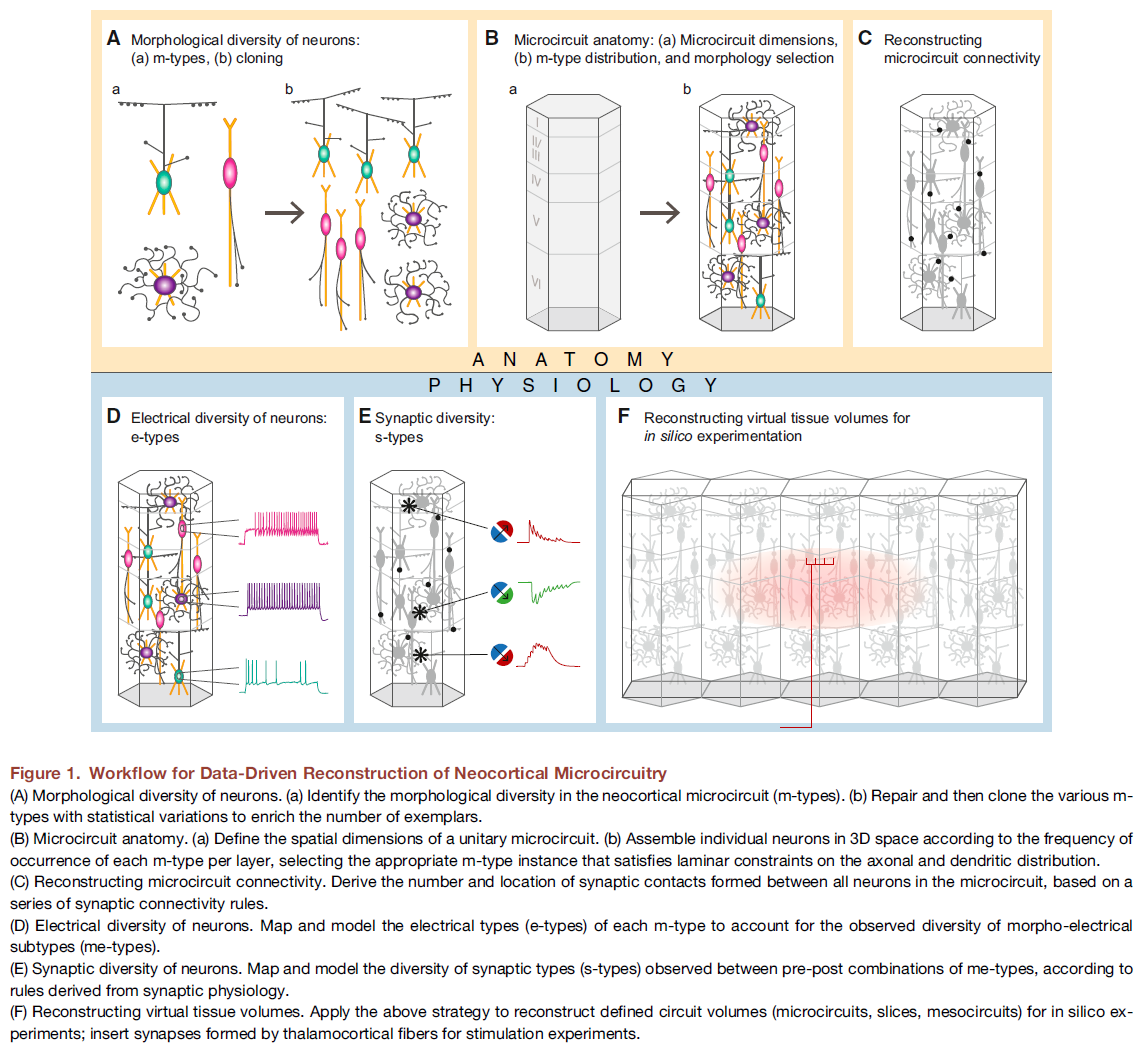
图 1. 数据驱动的新皮质微电路重建工作流程
(A) 神经元的形态多样性。(a) 确定新皮质微电路(m型)中的形态多样性。(b) 修复然后克隆具有统计变化的各种m型,以丰富样本的数量。
(B) 微电路解剖。(a) 定义单一微电路的空间维度。(b) 根据每层每个m型的出现频率在3D空间中组装单个神经元,选择满足轴突和树突分布的层流约束的适当m型实例。
(C) 重建微电路连接。根据一系列突触连接规则,推导微电路中所有神经元之间形成的突触接触的数量和位置。
(D) 神经元的电多样性。映射和建模每个m型的电类型(e型),以解释观察到的形态电亚型(me型)的多样性。
(E) 神经元的突触多样性。根据突触生理学得出的规则,映射和建模在me型的前后组合之间观察到的突触类型(s型)的多样性。
(F) 重建虚拟组织体积。应用上述策略为计算机实验重建定义的电路体积(微电路、切片、中间电路);插入由丘脑皮质纤维形成的突触用于刺激实验。
RESULTS
Neuron-type Nomenclature
神经元在大脑中的位置、形态、电特性、投射以及它们表达的基因和蛋白质方面有所不同(有关评论,请参见Harris and Shepherd, 2015; Markram et al., 2004)。这些特性的组合意味着神经元类型的巨大多样性。鉴于其他维度缺乏足够的数据,用于此初稿数字重建的神经元分类仅考虑了层、局部形态和电生理学。形态类型的命名基于上个世纪最常用的名称(Connors and Gutnick, 1990; DeFelipe, 1993; DeFelipe et al., 2013; Douglas and Martin, 2004; Fairén et al., 1984; Hestriand and Armstrong, 1996; Kawaguchi and Kubota, 1997; Kisvárday et al., 1985; Oberlaender et al., 2012; Somogyi et al., 1982, 1998; Svoboda et al., 1997; Szabadics et al., 2006),用层前缀扩展(例如,Layer_Morphology、L5_MC用于第5层Martinotti单元)。基于Petilla约定(Ascoli et al., 2008)的电类型被视为子类型(例如,非调节子类型的L5_MC_NAC;参见实验程序)。当有足够数量的投射神经元的全脑轴突追踪数据可用时(例如,L5_TTPC_CP和L5_TTPC_CT代表皮质脑桥和皮质顶盖亚型;Hallman et al., 1988; Wang and McCormick, 1993; 回顾,参见Ramaswamy and Markram, 2015),所提出的分类可以扩展到包括投影子类型。同样,当有足够的单细胞基因和蛋白质表达数据来系统识别细胞时,它可以扩展到包括分子亚型。图2中提供了每种m型使用的缩写。表1中提供了本研究中使用的命名法与文献中出现的替代名称之间的映射。
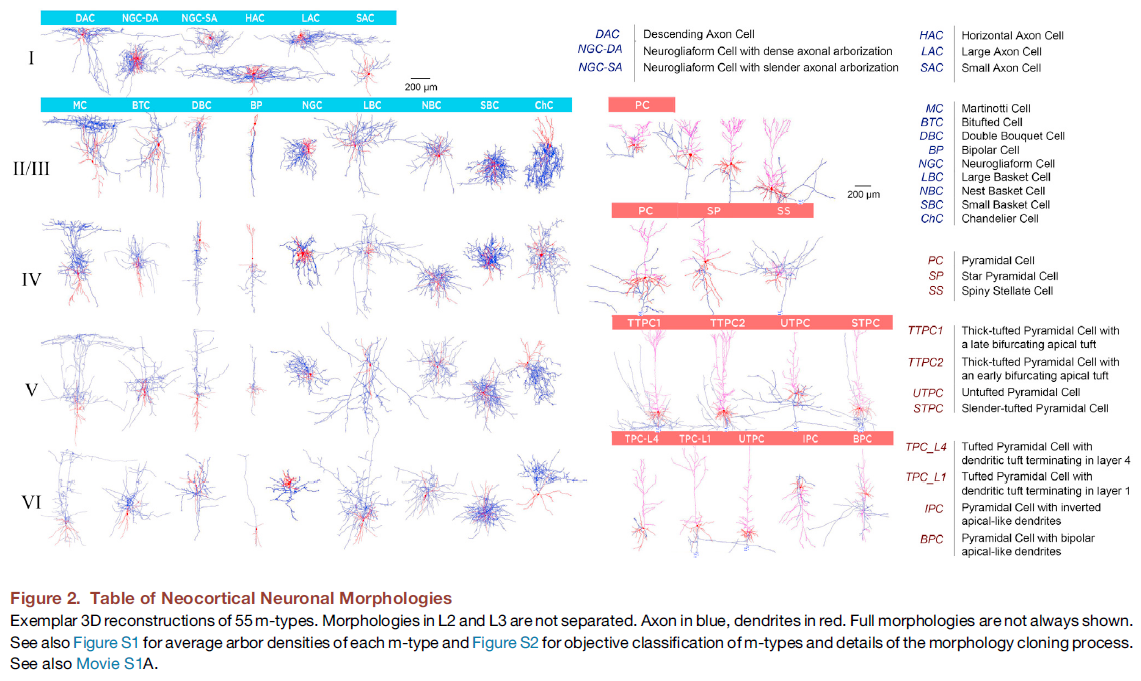
Morphological Diversity of Neocortical Neurons
我们在体外切片中使用膜片钳电极记录并标记了P14雄性Wistar (Han)大鼠体感皮层中所有六层的超过14000个神经元。在这些神经元中,有2052个被充分染色以允许专家根据其树突和轴突轴的公认特征将其分类为m型,这是由早期神经解剖学家发起并至今仍在使用的程序(Fairén et al., 1984; Karagiannis et al., 2009; Karube et al., 2004; Kawaguchi and Kubota, 1997; Kisvárday et al., 1985; Larkman, 1991a; Perrenoud et al., 2013; Peters and Kaiserman-Abramof, 1970; Ramón y Cajal, 1909, 1911; Somogyi et al., 1982, 1998; Wang et al., 2004; Yuste, 2005)。我们能够以数字方式重建其中1009个神经元的子集。这允许使用基于特征聚类的客观方法(见下文)验证专家分类,并提供重建微电路所需的数字神经元模型的初始池。在少数情况下,我们没有对已知存在于微电路中的稀有m型进行形态重建(L5_BP、L5_ChC、L6_NGC;Oláh et al., 2007; Szabadics et al., 2006)。这些是使用来自相邻层的相同形态的示例来表示的。尽管L6水平和亚板锥体细胞(L6_HPC和L6_SPC)存在于数据集中,并且在文献中也有报道(Ghosh and Shatz, 1993; Hevner et al., 2001),但染色的质量不够进行可靠的重建。这些形态在初稿中没有体现。
汇总文献中的形态重建和报告,我们区分了55个m型(如果单独考虑第2/3层,则为65,如果还考虑L6_HPC和L6_SPC,则为67;图2)。抑制类型主要通过轴突特征和兴奋类型通过树突特征来区分(评论见Markram et al., 2004; Ramaswamy and Markram, 2015; Spruston, 2008)。图S1显示了55种主要m型中的每一个的多个示例的叠加,图S2A和S2B说明了客观分类。虽然在某些情况下,可能能够在m型之间引入更精细的分离,但这会限制单个类型的样本大小,从而降低分类的可靠性。
除了第1层外,所有层都存在相同的抑制类型,第1层包含一组独特的抑制神经元类型。如L23_PC的多样性所示(图2,右上),锥体细胞形态在不同层(图2,右)和层内深度也不同。由其局部形态定义的锥体细胞类型的数量从上层到下层增加。几种类型的中间神经元(例如LBC和DBC)具有轴突轴,当它们位于上层时倾向于下降到更深层,当它们位于更深层时倾向于上升到上层。与这一趋势一致,一种锥体细胞(L6_IPC)也具有倒置的轴突轴。
使用从不同动物获得的每种m型的多个样本,我们开发了一种修复过程来恢复切片过程中切割的轴,并使用体内重建的神经元进行了验证(参见实验程序;Anwar et al., 2009)。为了生成更大的独特形态库,我们克隆了每个m型的多个示例(图S2C-S2F)、抖动的分支角度和克隆中的截面长度(参见实验程序)。针对从重建神经元获得的特征分布验证了所得群体的形态测量特性(参见实验程序)。这种方法使我们能够建立尊重生物变异性的神经元形态数据集(参见视频S1A)。可通过NMC门户获得用于修复和克隆体外神经元形态以及将神经元自动分类为55个m型的软件应用程序。
Reconstructing Neuron Densities, Ratios, and Composition
Positioning Morphologically Reconstructed Neurons
Morpho-Electrical Composition
Digital Reconstruction of Connectivity
Reconstructing Neuronal Physiology
Reconstructing Synaptic Physiology
Simulating Spontaneous Activity
Simulating Thalamic Activation of the Microcircuit
Reproducibility of Emergent Properties
Reproducing In Vivo Findings
Neuronal Responses to Single-Whisker Deflection
Anti-correlated Inhibitory Activity Cancels Out Highly Correlated Excitatory Activity
Temporally Sequential Structure during Spontaneous Activity of L5 Neurons
Soloists versus Choristers
Functional Implications
DISCUSSION
本文介绍了新皮质微电路的第一个数字重建草案,它集成了神经元形态、层高、神经元密度、兴奋性与抑制性神经元的比率、形态学和电形态学组成、电生理学以及突触解剖学和生理学的实验测量(参见"重建数据"和表S3)。它已针对重建中未使用的一系列单独的解剖学和生理学测量结果进行了验证(参见"验证数据"和表S3)。重建提供了对新皮质微电路的广泛解剖和生理特性的预测(Box 1)。重建的模拟显示了一系列紧急网络活动状态,从同步状态到异步状态急剧转变。在光谱的这一特定点上,数字重建再现了体内研究的许多发现,从而可以更深入地研究其潜在的细胞和突触机制。它们还可以进行迄今为止在体外或体内不可能进行的实验。对重现微电路关键功能特性所需的网络大小的研究表明,它大致相当于用作重建基础的新皮质组织的体积。这证明这种规模的网络是新皮层信息处理所需的最小功能单元。
Validity of the Digital Reconstruction
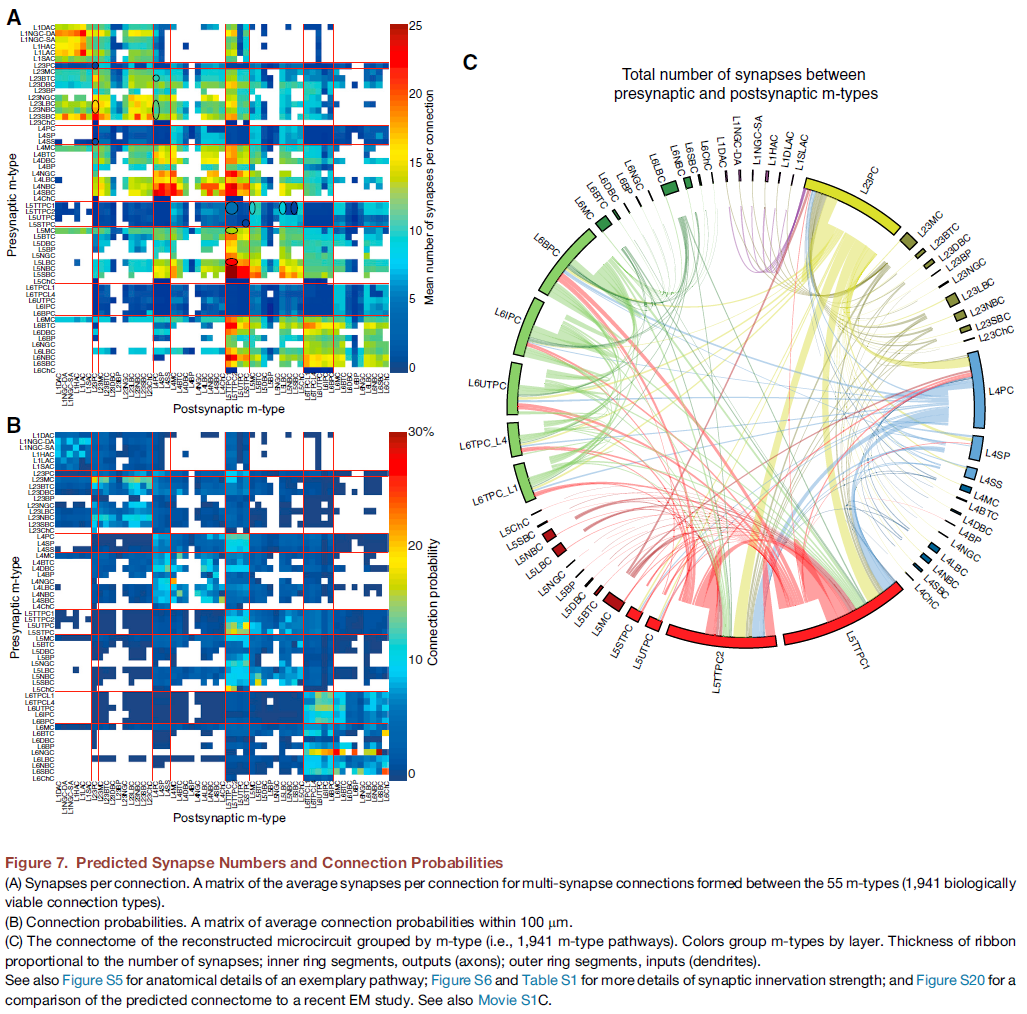
重建当然包括由于实验数据集的错误和差距以及对生物学原理的不完全理解而导致的错误。例如,额外的细胞类型标记将提高形态组成的准确性,饱和的EM重建可用于进一步验证派生的连通性,更多报告突触反应的电压和电流组合测量的实验将加强关于量子电导和连接特异性突触动力学的结论 , 并进一步表征不同突触对[Ca2+]o的敏感性,可以更准确地划分不同活动状态之间的转换,并可以更精确地确定每个神经元和突触类型在维持和转换状态中所起的作用。例如,最近的一项EM研究发现,每个连接的突触数量比对Peters规则的幼稚解释所预测的要多(Kasthuri et al., 2015)。将相同的分析应用于数字重建产生了类似的结果(图S20在计算机上与图7D、7F和S6B (Kasthuri et al., 2015))。由于优先修剪具有少量突触的连接,这些特性出现在数字重建中(Reimann et al., 2015)。
在重建过程的多个阶段进行的验证测试降低了错误可能导致重建或模拟其紧急行为时出现重大错误的风险。例如,针对独立数据验证电神经元模型可以将网络的紧急行为与我们对离子通道动力学和分布的有限知识的影响隔离开来。更一般地说,重建通过了广泛验证其潜在解剖学和生理学的多项测试。例如,细胞形态、密度、组成和连接性方面的重大错误将难以重现在12块实验中发现的神经元组装类型、锥体体上GABA能突触的数量、蛋白质染色模式、逐层突触密度、连接概率、布顿密度和分布等(见表S3)。这些特性很好地位于实验报告的范围内。使用来自不同动物的数据并结合统计变化在多次重建中观察和预测的可重复性提供了证据,证明它们是稳健的。
尽管据我们所知,重建是迄今为止最详细的,但它忽略了微电路结构和功能的许多重要细节,例如间隙连接、受体、神经胶质、脉管系统、神经调节、可塑性和体内平衡。此外,它代表了一只雄性大鼠在年轻时仅一个大脑区域的快照。这限制了可以得出的结论的普遍性。例如,在用于研究的年龄的动物中,树突形态已经成熟到成人水平(Larkman, 1991a; Romand et al., 2011),但上行轴突可能没有完全代表,当然也没有完全成熟(Romand et al., 2011)。然而,在更高水平的生物学细节(例如,包括神经胶质、受体和信号通路)的研究以及对不同年龄、性别和物种的动物以及疾病模型中不同大脑区域的研究,可以使用重建作为一个参考点。与重建一致的调查结果将表明重建过程中使用的组织原则是充分的;差异可能指向新的原则。例如,如果将连接算法应用于另一个大脑区域或不同年龄或属于不同物种的动物未能产生与实验结果一致的结果,这将指向连接规则的特定变化。
验证失败也可能表明实验数据存在错误。例如,重建表明之前十几项研究的细胞密度都太低,无法解释脊柱和突触的密度,这表明新的实验验证了这一预测。重建还表明,许多实验低估了突触电导,并表明不考虑bath中钙水平的体外实验可能会误解他们的发现与体内条件的相关性。这些示例说明了重建过程如何不采用表面价值的实验数据,而是尽可能使用互补的相关数据集来限制作为参数的使用。
Functional Implications
对自发和诱发活动的模拟解释了不同类型突触对Ca2+的不同敏感性,并探索了Ca2+水平的变化,揭示了从同步到异步行为的一系列活动状态。改变Ca2+水平会深刻地改变整体E-I平衡,从而改变网络沿光谱的位置,从而导致活动状态之间的急剧转变。这些计算机预测已通过新的体外实验得到验证。
进一步的模拟表明,发生转变的Ca2+水平在使用来自不同动物的数据的数字重建中有所不同,这在神经元脉冲和网络空间分辨率的变化中占很大比例。我们还发现,在个体重建中对Ca2+水平(~0.05 mM)进行小幅调整会显著降低其生理变异性。这些模拟提供了一个例子,说明个体神经解剖学的变化如何导致功能差异。
受这一发现的启发,我们进行了进一步的模拟,结果表明激活或抑制特定层、神经元和突触连接也会沿着频谱改变网络。虽然从之前的理论发现中众所周知,改变E-I平衡会改变网络的状态(Brunel, 2000; van Vreeswijk and Sompolinsky, 1996),但模拟进一步表明,任何会对不同类型突触的突触动力学产生不同影响的机制(例如,通过神经调节;有关评论,请参见Lee and Dan, 2012; Zagha and McCormick, 2014)都能够以复杂的方式改变活动机制之间的界限。我们推测,数字重建无法再现的其他紧急特性,例如具有两个亚稳态固定点的UP和DOWN状态(Steriade et al., 1993),可能需要丘脑-皮质相互作用(Hughes et al., 2002),皮质-皮质相互作用(Timofeev et al., 2000),内在振荡器(Lörincz et al., 2015; Sanchez-Vives and McCormick, 2000), 或神经调节(Constantinople and Bruno, 2011; Lörincz et al., 2015; Sigalas et al., 2015)。因此,细胞或突触生理学的调节可以作为动态重新配置网络以满足不同计算要求的机制。
Reproducing In Vivo Findings
尽管数字重建主要基于体外数据,并非旨在重现任何特定实验或捕捉复杂的体内条件,但它产生的结果在质量上与许多主要的体内发现相当,并且在这些实验中做出了超出可能的预测,而无需调整任何模型参数。
例如,数字重建使解决一个长期存在的问题成为可能,该问题涉及在体内实验中经常观察到的不相关活动背后的机制(Haider et al., 2006)。先前的理论工作表明,不相关的活动可能是紧密相关的兴奋电导的结果,这些兴奋电导被反相关的抑制电导有效抵消(Renart et al., 2010; van Vreeswijk and Sompolinsky, 1996)。我们的模拟使用并非专门设计用于解决这个问题的模型,证实了这种效应是网络的一种新兴属性。模拟进一步表明,体内的皮层活动接近同步异步频谱的关键转变,超过该转变的再生活动导致神经元雪崩(另见Beggs and Plenz, 2003)。围绕这一转变,脉冲活动与突触输入中的精细时间结构高度相关,反映在兴奋性和抑制性电导之间的短暂失衡中。空间隔离输入之间的最大区分、精细时间结构(如三元)的生成以及类似独奏者和合唱团的行为都出现在过渡附近。最近的一项研究通过实验描述了维持网络接近这种转变的可塑性机制(Delattre et al., 2015)。
重现这些体内发现令人惊讶,因为数字重建是基于从未成熟大鼠体感皮层获得的数据和结构原理,而许多体内发现来自成年动物的不同新皮质区域,有时属于其他物种。重建再现这些现象的事实表明它们来自新皮质微电路的基本特性。
Concluding Remarks
这项研究表明,原则上可以重建新皮质微电路结构和功能的综合视图,使用稀疏的互补数据集来预测尚未通过实验测量的生物参数。虽然目前的数字重建已经可以用来深入了解微电路的运作方式,但这只是第一步。为了促进新实验数据的整合和对其所依据的原则的挑战,我们创建了一个公共网络资源,提供对重建中使用的实验数据、模型和工具的访问(新皮质微电路协作[NMC]门户,https://bbp.epfl.ch/nmc-portal;Ramaswamy et al., 2015)。这将允许社区整合他们自己的数据,执行他们自己的分析,并测试他们自己的假设。
EXPERIMENTAL PROCEDURES
补充实验程序中提供了详细说明。
Data Acquisition
Neuron Morphology
在对300毫米厚的脑切片进行全细胞膜片钳记录后,从幼年大鼠后肢体感皮层的生物细胞素染色神经元的数字3D重建中获得神经元形态(Markramet al., 1997)。在一些重建的神经元中,bouton位置被注释在轴突上(Wang et al., 2002)。重建使用Neurolucida系统(MicroBrightField)。
Neuron Electrophysiology
Synaptic Anatomy
Synaptic Physiology
Tissue Immunohistochemistry
Electron Microscopy
Multi-electrode Array Experiments
Manipulating [Ca2+]o
Reconstruction Process
Digital Neuron Morphologies
在3D重建之后,使用修复算法(Anwar et al., 2009)恢复神经元形态的切割端。神经元轴被数字解开以补偿由收缩引起的曲折,并且克隆了神经元形态(参见补充实验程序)。
Electrical Neuron Models
使用多达13种活性离子通道类型和细胞内Ca2+动力学模型生成基于多室电导的神经元模型。轴突初始节段(AIS)、胞体、基底树突和顶端树突被分离。中间神经元仅包含一个树突区域。每个区域都收到一组单独的频道(参见NMC门户,https://bbp.epfl.ch/nmc-portal;Ramaswamy et al., 2015)。在轴突中,仅模拟了AIS。每个AIS由两个固定长度的部分表示,每个部分的长度为30 毫米。AIS直径是从用于模型拟合的重建形态中获得的。在AIS中检测到的动作电位被发送到突触后突触,延迟对应于轴突长度,假设轴突速度为0.3 m/s。如前所述(Druckmann et al., 2007),使用基于特征的多目标优化方法拟合神经元模型。
The Microcircuit Volume
Cellular Composition
Synaptic Anatomy
Synaptic Physiology
Thalamic Innervation
Simulation
Microcircuit Simulation
重建的微电路使用NEURON仿真包进行仿真,增强了在超级计算机上的执行(Hines and Carnevale, 1997; Hines et al., 2008a, 2008b),以及用于处理微电路的设置和配置以及结果的输出。
In Silico Experiments
通过模拟神经元胞体的电流注入来实现去极化。电流表示为每个神经元的第一个脉冲阈值的百分比。通过根据特定m型的三个曲线改变突触传递的使用参数来模拟[Ca2+]o的变化(见图S15)。通过用胞体电流注射(100%阈值)使目标群体超极化来进行神经元计算机敲除实验。丘脑纤维刺激在小柱的圆形簇上进行。用于复制先前体内实验的方法在补充实验程序中进行了描述。
Data Analysis
Supercomputing
Visualization
Software Development
SUPPLEMENTAL INFORMATION