郑重声明:原文参见标题,如有侵权,请联系作者,将会撤销发布!

Trends in Neurosciences, no. 5 (2010): 220-229
Abstract
回合空间记忆均涉及海马体神经元回路中复杂联想的编码。这种记忆轨迹可以通过巩固过程从短期到长期稳定下来,这种巩固过程涉及睡眠和休息期间原始网络发放模式的“重新激活”。清醒经验能够被许多不同的大脑区域回放,但海马体的重要作用在“重新激活”过程的组织中。越来越多的证据表明,海马体中的尖波/波纹(SWR)事件可以协调记忆轨迹的重新激活并指导其在皮质回路中的恢复。尽管机制仍不确定,但越来越多的共识认为,这种SWR指导的全脑记忆轨迹的重新激活可能是记忆巩固的基础。
Box 1. Hippocampal firing patterns during exploration and sleep
清醒探索和休息期伴随着局部场电势(LFP)的不同模式。在机警的清醒和探索过程中,海马体LFP的特征是θ波段振荡(5–12 Hz,图1a)(参考文献[94])。在啮齿动物中,睡眠可大致分为快速眼动(REM)(再次以θ振荡为特征)和non-REM epoch。Non-REM睡眠以SWR事件为主(图Ia,b)。SWR的两个成分是CA1辐射层(stratum radiatum)中的负“尖波”[95]和CA1锥体层中的瞬态快速“波纹振荡”(150-250 Hz)[16,21]。这些事件是由CA3细胞的同步发放引发的,依次促进了波纹振荡和CA1细胞的同步发放(相对于θ epoch,同步增加了5倍)(参考文献[17,27,96,97])。尽管CA1神经元和锥体细胞的发放率都有显著增加,但锥体细胞的发放增加更大[97]。反过来,这些事件导致内嗅皮层和下丘脑的去极化和网络活动的增加[91,98],以及跨多个大脑区域的协同发放。尽管主要通过啮齿动物研究来表征,但是在灵长类和人类中也已发现了SWR活动,尽管频率较慢[99,100]。

Box 2. Outstanding questions
- 尽管研究表明重新激活的模式与动物的先验行为有关,并且SWR的发放可以促进空间学习,但尚无研究表明重新激活本身是反映获得的记忆轨迹还是在巩固过程中起作用。因此,必须问:
- 具体来说,在复杂的记忆任务中,重新激活是否反映了已学到的内容?
- 例如,在一项必须学习许多项目的任务中,与学得不好的项目相比,被召回项目的表示是否被更强烈或更频繁地重新激活?
- 通常,重新激活可以预测对所获取信息的未来召回吗?
- 尚无有关重新激活如何促进巩固的数据;重新激活是否可能导致海马体信息的转移,和/或加强不同大脑区域的助记项之间的关联?
- 重新激活是否在巩固所有类型的海马体依赖性记忆或仅某些类型的记忆中发挥作用?
- 另一个未解决的问题与重新激活背后的机制有关。虽然可能需要突触可塑性来存储重新激活的模式,但尚不清楚哪种网络事件会导致可塑性变化以及在细胞和突触水平上会发生什么变化。
- 最后,还不清楚控制特定模式重新激活的因素。海马体外输入是否控制重新激活哪些模式?
The hippocampus and consolidation
学习和记忆过程对于生物体使其行为适应不断变化的环境,使其能够利用先前的经验来预测未来至关重要。建立位置和事件的稳定记忆并不是一个单一的过程。最初,回合记忆是不稳定的,并且在持续的经验中很容易退化。随着时间的流逝,这样的记忆会抵抗干扰的破坏性影响,最终可以持续数十年。理解这些过程背后的神经机制仍然是神经科学的重要目标。
自从观察到海马体受损患者在形成新记忆方面出现缺陷[1]以来,许多研究表明,海马体在记忆处理的所有阶段(即编码,巩固和记忆)都参与了回合空间记忆的形成[2–6]。目前的理论认为,在初始编码后,海马体具有时间限制的作用,其中记忆轨迹向永久性新皮层存储的转移与海马体脱离有关[5-7],或者,海马体永远不会脱离,记忆恢复将继续与原始经验的海马体和皮层表征的激活有关[8,9]。无论如何,新编码的信息通过涉及海马体-皮层交互的巩固过程得以稳定。
在人类和啮齿动物上进行的多项研究表明,睡眠可能在这种系统级巩固中起作用[10-13]。但是,由于睡眠和记忆都不是单一现象,因此对这些过程的理解变得很复杂,前者由多个系统组成[14],而后者则由多个阶段组成[15]。来自啮齿动物的越来越多的证据表明,称为尖波/波纹(SWR,Box1)的网络事件在海马体依赖性记忆巩固中起着关键作用。这些SWR最明显地出现在休息和睡眠间隔中,与快速的眼球运动无关(non-REM睡眠),这些时期代表了海马体神经元之间最高发放同步的时间,从而为海马体内部的可塑性以及海马体与其他大脑结构之间的交流提供了条件[16-18]。
海马体依赖性记忆形成可能在两个主要阶段发生。首先,海马体在清醒时快速地编码记忆。然后,在“离线时段”内,海马体会“重新激活”记忆轨迹,从而促进转移到皮层网络中的长期储存状态,或促进海马体内外不同大脑区域的轨迹之间的关联[18,19]。
许多小组都观察到了SWR期间清醒神经活动的重新激活。在这里,我们回顾了在啮齿动物中使用单独单位记录的大量工作,这些工作证明了海马体和其他大脑区域的重新激活,并评估了在记忆环境中放置此现象的进展。通过这样做,我们讨论了重新激活的模式如何反映动物的行为。我们还总结了指向跨大脑区域协同重新激活的数据。
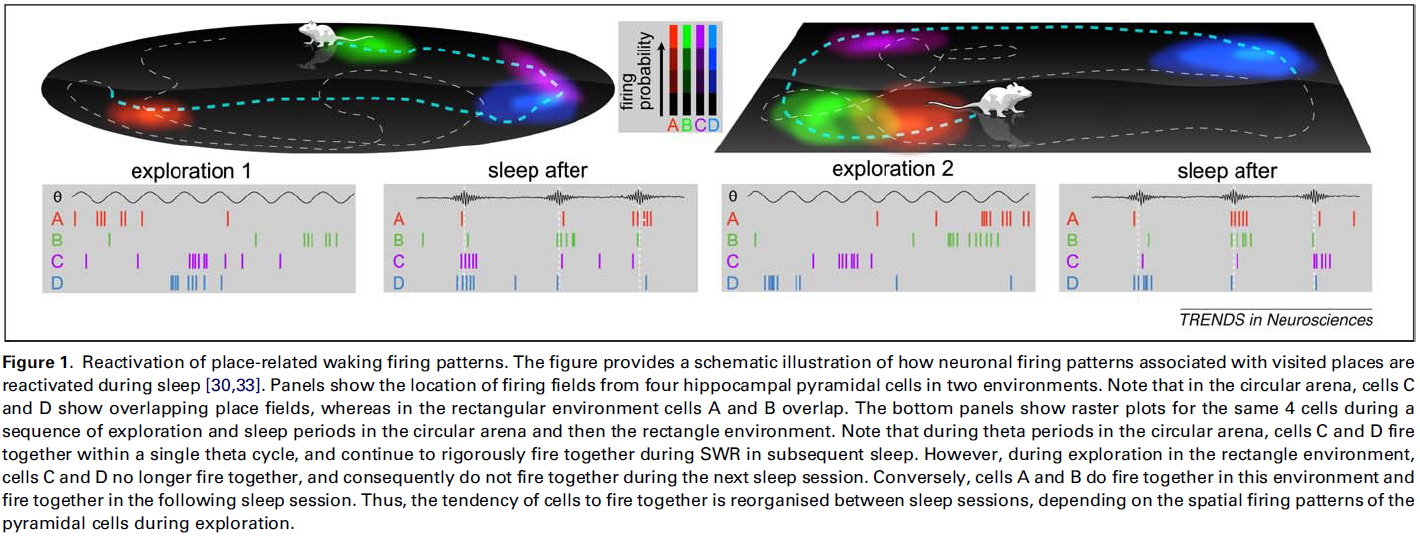
Reactivation of waking sharp wave/ripple (SWR) firing patterns during sleep
CA1和CA3区域中兴奋性神经元的发放模式在活跃的清醒和休息时段的网络振荡过程中表现出特征差异 (Box 1),并且据认为在记忆处理中起着不同的作用。在探索过程中,每个细胞都显示出稀疏的发放,这种发放倾向于被限制在环境的离散区域,称为“位置场”[20,21] (Box 1,图1c)。这些“位置细胞”的子集在不同的位置发放,共同构成重叠场的拼凑,以覆盖整个环境 (Box 1,Figure Id)。作为结果,根据动物的行为及时组织了位置细胞组的发放方式。也就是说,随着动物的探索,不同组的细胞在环境中的特定位置一起发放。保留这些发放模式的时空组织可以为清醒经验的记忆轨迹提供基础。CA3区域是这种轨迹的一个潜在存储库,该区域具有强烈的循环连接性和高度可修改的突触[22,23]。在这里,联合发放细胞的组可以通过突触权重的变化形成关联。此后,仅通过这些细胞的一个子集发放将倾向于通过它们的循环连接组成整个组(或“集合”),从而召回整个“模式”[23-25]。在这样的网络中,当某些细胞(“启动程序细胞”[18])的同步激活触发整个组装模式的完成时,先前存储的模式可能会自动重复出现[26]。因此,CA3区的模式完成可以促进休息和睡眠期间清醒模式的回放。
CA1和CA3区域中偶发的同步发放突发事件标志着不动性和non-REM睡眠,并与SWR事件协调。这些事件被认为是在CA3区域内引发的[17,27],因此SWR期间的整体活动主要受CA3区域内以及CA3和CA1区域之间已建立的突触权重的支配。因此,在SWR期间的发放模式可以通过模式完成来成形,从而促进代表记忆轨迹的海马体组件的联合发放。反过来,像排练一样的“重新激活”存储的记忆轨迹可以促进它们的巩固[28]。
表现出啮齿动物行为的大规模多通道记录技术的发展[29]提供了支持这一推测的生理学证据。使用这些技术,实验者可以在动物探索和睡觉时记录其大细胞集合的发放模式,从而有可能测试在随后的睡眠中是否重新激活了位置细胞的清醒活动模式。首先由Wilson和McNaughton展示的,具有重叠位置场的CA1锥体细胞在探索后的睡眠过程中比编码不同位置的细胞具有更大的共同发放倾向。而且,这种关系在探索之前的睡眠中已大大降低[30]。因此,根据介入探索中位置场的配置,不同组的细胞从一个睡眠到另一个睡眠一起发放[31-33](图1)。重要的是,在SWR期间,睡眠重新激活最强[31,33,34]。这些技术已广泛用于证明利用不同类型的分析如何在睡眠SWR中反映清醒活动的模式[35]。下一步重要的工作是,表明重新激活的模式所反映的不仅仅是在先前探索的环境中的位置场的配置。

Reactivated hippocampal firing patterns reflect exploratory behaviour
如果海马体发放模式的重新激活是海马体依赖性记忆巩固的基础,那么重新激活的模式不仅应反映动物探索的环境,而且还应反映其近期行为。多项研究表明,睡眠发放模式反映了动物在探索过程中采取的路径。当动物在狭窄的轨道上奔跑觅食时,位置细胞以受控的顺序发放,而动物则依次穿过每个位置场。在这些条件下,大多数位置细胞倾向于仅向一个方向发放,因此这些细胞对中的联合发放模式表现出时序偏差(即一个细胞倾向于在另一个细胞之前发放)。在探索过程中,这种时序偏差会保留到随后的睡眠中[36]。此外,进一步的研究表明,在睡眠SWR之后,在轨迹运行中回放了活跃细胞的整个发放序列,但是比轨迹运行时的时间尺度更快[37,38](如图2所示)。应该注意的是,这种序列回放也在REM睡眠期间发生[39]。
SWR在休息期间的全体活动也反映了动物在“开放领域”环境中的行为,在这种情况下,动物可以自由探索运动场,而不必遵循刻板印象。在这里,当动物花了更长的时间探索记录运动场时,在SWR期间清醒的发放模式会更可靠地重新激活[32]。此外,如果动物在路径上来回穿梭花费更多的时间,则即使在线性轨道上,重新激活也会得到改进[40],这总体上说明重新激活会随着经验的增加而提高。
SWR发放模式不仅反映了环境,而且反映了行为,这进一步表明来自以下事实:在以后的睡眠中,访问频率较高的地方会更强烈地重新激活。结果表明,在随后的睡眠过程中,编码特定位置的细胞的发放同步性随着在先前探索期间在该位置花费的时间而增加[33]。因此,重新激活的模式偏向访问量最大的地方(图3)。
总之,这些发现表明,与探索有关的发放模式在睡眠期间会重新激活。所有这些研究的警告是,在这些实验中没有执行特定的学习范式。最近的工作表明,学习后抑制CA3锥体神经元突触释放既抑制了CA1区域的重新激活,又抑制了上下文相关记忆的巩固,表明这些现象是相关的[34]。将重新激活与记忆联系起来的其他证据来自这样的事实,即记忆受损的老年大鼠表现出受损的序列重新激活[41]。最后,最近的两个报告表明,休息期间的SWR释放与巩固有关。通过实时检测SWR,有可能在SWR开始时刺激传入CA1区,从而阻止SWR的完整表达。在执行空间记忆任务后休息期间SWR中断的大鼠在几天内学习能力较弱[42,43]。这些数据表明SWR在空间记忆巩固中的作用。但是,在这些实验中,SWR期间的电刺激有可能改变了CA3-CA1突触的连接,从而破坏了学习[44,45]。此外,还不清楚单独的SWR还是SWR期间的重新激活是否会促进巩固。因此,到目前为止,尚未直接显示,重新激活的模式表示巩固过程中的记忆轨迹。即便如此,我们知道,重新激活的模式代表了以前的清醒经验,首先需要将其存储在海马体中。下节将讨论这些过程的潜在机制。
Does synaptic plasticity underlie reactivated patterns?
Donald Hebb是第一个推测记忆可以由细胞群体协调“回荡”活动编码的,并且由于这个活动的“持久性或重复性”,这种活动数据包可能被稳定化为更永久的结构(赫布细胞组装)。这将倾向于诱发“持久的细胞变化,从而增加其稳定性”[26]。长期增强(LTP)的发现提供了潜在的生理机制[46],并且LTP可以通过突触前和突触后动作电位的频繁配对(即Hebbian学习规则)来诱导。多种形式的突触可塑性取决于NMDA型谷氨酸受体(NMDAR),包括CA3和CA3–CA1区域锥体细胞之间突触处的LTP[47,48]。已经表明,阻断NMDAR会损害空间学习[49,50]和某些海马体依赖性记忆的巩固[51]。因此,依赖于NMDAR的突触可塑性可以作为后续重新激活的记忆轨迹的存储基础。
Spike timing dependent plasticity (STPD) and sequence replay
探索一维(1D)环境后位置细胞序列的回放可以通过在轨迹运行过程中位置细胞之间形成不对称关联来解释,这样在睡眠期间,细胞倾向于以与探索期间相同的顺序发放。这些不对称关联可以按照一种称为STDP的Hebbian机制的形式建立。在此依赖于NMDAR的过程中,仅当突触前细胞在突触后细胞之前发放时才发生增强。如果突触后细胞先被发放,则导致抑制[52,53]。在轨迹运行过程中,位置场相对于动物奔跑的方向反向扩展,该过程取决于NMDAR[54-56]。可以通过在细胞与相邻位置场之间形成更强的关联来解释这种效果,从而使每个细胞都增强了与动物运动方向上紧挨着的突触后细胞对的连接权重。这些不对称的关联可能会使细胞偏向于在下一条轨道上更早地发放,并可能有助于形成组装序列以用于随后的重新激活。
Synaptic plasticity in the reactivation of places
相反,在2D环境中,动物在探索环境时可以采取许多不同的路径。在这里,具有重叠场的细胞对不一定会按照设定的前后顺序发放,因为动物不会始终如一地越过一个场。即使细胞在STDP所需的清醒过程中没有表现出时序偏差,也会发生位置的重新激活[32]。细胞的发放率与清醒和睡眠时段之间相关,并且睡眠时段之间频率的变化可以促进不同细胞亚群的联合发放[57,58]。但是,仅凭这种“频率重新激活”不能解释在重新激活过程中观察到的细胞共同发放[33],并且可能需要涉及细胞之间突触可塑性的相关变化。最近的一份报告表明,在随后的睡眠中,细胞的联合发放趋势(共同发放)取决于它们在探索过程中一起发放的次数。此外,这些细胞必须在较短的时间间隔(<50 ms)内一起发放,以观察重新激活[33],这让人联想到赫布学习规则所要求的协同性。实际上,最近的数据表明,不需要精确的前后顺序来诱导CA3–CA1区突触中的LTP[59,60],这表明LTP可能发生在反复共同发放的细胞之间的突触中,即使它们不按设定的时序发放也是如此。
将新的清醒经验的不稳定表示整合到先前存在的表示中,可能需要其他过程来防止其因干扰现有的表示而退化。

Reactivation after exploration of novel environments
空间学习通常涉及形成新遇到的环境的内部表示。新位置表示的稳定性需要依赖于NMDAR的可塑性。NMDAR封锁不会阻止代表熟悉环境的位置图的召回,也不会阻止在进入新环境后出现新的特定于上下文的位置图。但是,如果在经历新环境之前先进行管理,NMDAR封锁会在24小时后阻止重新建立这些新建立的位置图[61],这意味着编码和/或巩固被破坏,而不是检索过程被破坏。因此,在新环境中,海马体内的突触可塑性可以被上调,从而有助于对新的场所和事件进行编码。Tulving及其同事提出了“新颖性编码假设”,即长期存储信息的可能性随信息的新颖性而直接变化[62]。以这种方式,在新的经验之后增强的重新激活可以优先促进代表那些事件的记忆轨迹的巩固。与此假设相符,重新激活在新颖的探索之后比熟悉的环境更强[33,63](图3)。探索新环境与提高发放率[64,65]和协调CA1锥体细胞[66]有关,这可能会促进神经元的可塑性。此外,暴露于新环境会导致多巴胺能D1/D5受体激活促进LTP诱导的CA1阈值降低[67]。此外,接触新颖性会促进海马体乙酰胆碱[68]和去甲肾上腺素[69]的升高,这两者均可促进LTP[59,70]。
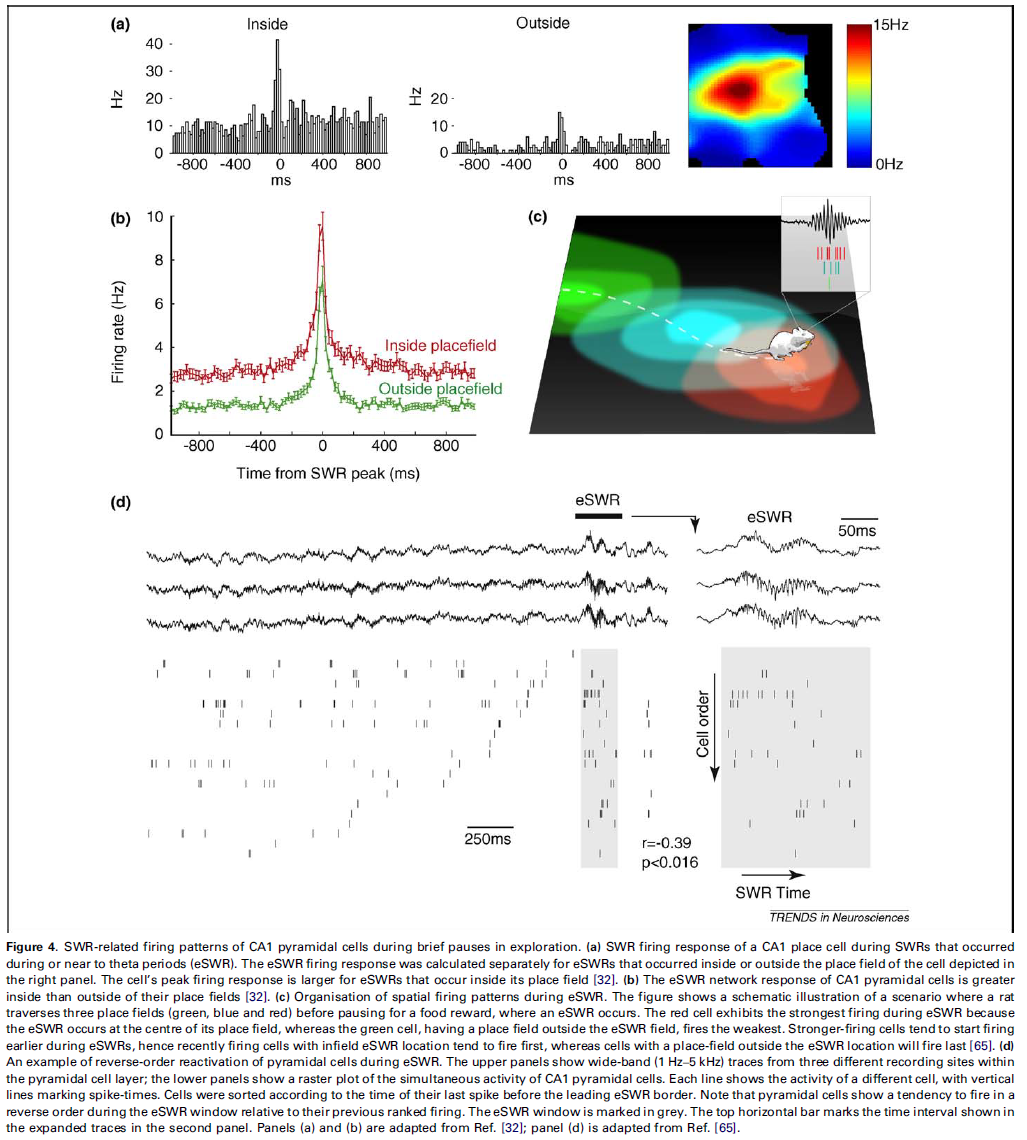
Plasticity during exploration-associated SWRs
SWR释放不限于长时间不动的睡眠或清醒时间。与探索相关的驻波比(eSWR)可能发生在θ振荡边界处的运动暂停期间,甚至是动物仍在移动时[32,66]。
CA1和CA3锥体细胞的发放模式表现出减少的位置选择性发放,这是θ振荡后经过的时间的函数[32,71],因此在长时间不动期间与SWR相关的发放受动物位置的影响较小[32]。相比之下,在eSWR期间,其所在区域内的细胞比外部细胞具有更强的发放能力(图4a)。这种增强的发放表示空间输入(可能源自内嗅皮层[72,73])与CA3区域的SWR相关发放重叠。这两个输入的组合产生了大于总和的CA1区域锥体细胞[32](图4b)。因此,在这些事件期间,在发放eSWR的位置上具有位置场的细胞会强烈发放,这为进一步加强它们之间的关联提供了条件[32]。因此,空间输入选择在eSWR期间激活哪些CA3和CA1细胞组件。一些研究报告说,在新环境中,类似于eSWR的高频振荡的发生率可能会比在熟悉的环境中更高,这表明这些现象与新环境相关的模式的重新激活有关[66]。这些数据表明SWR的双重作用,既有利于主动探索过程中的初始轨迹采集,又有助于促进存储轨迹的回放。实际上,最近的数据表明,海马体可以在编码和回放模式之间快速切换。
Reactivation during brief pauses in exploration
在记忆形成的两阶段模型中,一个假设是在睡眠期间会发生轨迹重新激活,以减少正在进行的轨迹形成所造成的干扰。但是,最近的发现表明,SWR在短暂的探索停顿期间也发生了重新激活[63,74,75]。
整体发放在奖励位置的SWR期间组织为序列(图4c)[63,65,73,75]。 第一次展示是在记录在线性轨道两端奖励位置的SWR期间进行的。在这些SWR中,位置细胞按顺序发放,以反映其位置场在轨道上的位置。但是,这种轨迹运行的“回放”顺序是相反的,因此,最近访问过的位置会被首先回放[74,75]。随后显示,在这些SWR中可以同时进行前向和反向回放,因此提出回放顺序取决于动物是即将离开食物奖励位置(前向回放)还是最近到达(反向)[74],尽管仍有待确认(但请参阅参考资料[76])。如上所述,在运动过程中不太可能通过不对称的突触修改来建立以相反顺序回放的序列,因为如上所述,STDP应该导致以与轨迹运行过程中相同的顺序(即前向)发放回放。在这种情况下,前向回放可能类似于在睡眠中看到的回放,反映出在探索过程中位置细胞之间形成的不对称关联。另一方面,反向回放很可能是由SWR之前的最近网络活动驱动的[65]。当位置选择驱动本身可以促进反向回放时,这种回放很可能在eSWR期间发生[65](图4c)。这一想法得到了以下事实的支持:在开放领域探索中,动物可以遵循不同的轨迹,在eSWR期间发放会显示最近活动的反向回放(图4d),而在SWR期间,在较长的固定时间内没有观察到反向回放[65]。此外,在eSWR期间几乎没有预期的开放领域探索的回放[65]。实际上,一些反向回放模型假设最近发放的细胞在SWR位置处的位置场增加了去极化[74,75,77]。
因此,在清醒活动期间,海马体可以在编码和回放模式之间快速切换。在探索过程中,反向回放可能有助于协调近期的脉冲活动,并可能促进表示环境中显著特征(例如奖励)的细胞之间的关联。在一维环境中,前向回放可能代表巩固的早期阶段。一些前向回放可能表示预期的上下文适当的检索,该检索为后续的路由准备做好了准备,例如在决策点处发生的连续位置的基于θ的前向扫描中[78]。这样的“检索”与巩固过程并不互斥。
Reactivation occurs throughout the brain
尽管海马体是否是空间/回合类记忆的最终存放处仍存在争议,但海马体与皮质之间的联合激活仍然是系统巩固概念的核心。有几条证据表明,整个大脑的重新激活是协调的。最初,Qin et al.研究表明,在海马体和顶叶皮层中,在行为过程中表达联合活动的成对细胞在随后的睡眠中继续共同发放[79]。这种重新激活发生在两个区域之间以及每个区域内,这表明重新激活是一种全局且协调的现象。这一发现已经通过研究得到扩展,该研究表明在来自几个不同皮质区域的神经元组之间发生了类似的重新激活,这些神经元不一定是单突触连接的[80]。这种协调且全脑范围的激活,与分布在不同大脑区域的记忆轨迹回放一致,且每个区域都贡献了轨迹的一部分,反映了其在清醒过程中的作用。Pennartz及其同事支持这一观点,报告说,与奖励相关的神经元,基底节清醒活动的重新激活最强,并且在睡眠期间海马体-纹状体整合在一起重新激活,这暗示了巩固位置-奖励关联的推定机制[81,82]。此外,最近从海马体和视觉皮层的记录中获得的数据表明,行为相关序列的连贯重新激活趋于在两种结构中同时发生[37]。使用如上所述的类似范例,在轨迹运行过程中建立海马体细胞序列。然而,一些视觉皮层细胞还显示出发放受限于轨道的各个部分(可能是相对于视觉信号的发放),因此也以固定顺序发放。在慢波睡眠过程中,视觉皮层和海马体细胞中这些序列的片段均被回放。尽管此序列回放很少,但它们倾向于同时发生。最后,最近的数据表明在海马体SWR期间,前额叶皮层的激活优先发生[83]。
Hippocampal–neocortical dialogue during sleep
在上一节中,我们描述了海马体和其他皮质区域之间如何协调重新激活。但是,仍然缺乏对这种协调回放背后的生理机制的描述。海马体和新皮层之间的网络交互已被详细检查。这项工作大部分旨在描述海马体SWR与新皮层网络振荡的时间关系。研究表明,驻波比在时间上与皮层纺锤体(10–18 Hz),慢波振荡和慢波相关的三角波[84–87]保持一致,这些确实可能有利于海马体-脑皮质对话。海马体网络模式与新皮层缓慢振荡的同步已引起特别关注。新皮层的缓慢振荡与上下状态有关:在慢波睡眠和麻醉过程中,新皮层神经元的膜电位显示出双峰分布,并且在增加的发放块中称为发放脉冲(称为"向上状态"),随后处于静止期("向下状态")[88]。一些研究表明,SWR的活动受皮层上下状态的调节,尽管这些研究的结果并不一致。一些研究表明,SWR倾向于在向上状态下发生[37,84,86],而另一些研究则表明,它们优先发生在从下到上的状态转变期间[89,90]。这些差异可能是由于用于估计上下状态发生的方法不同(大脑区域不同)所致。一些研究表明,SWR期间的重新激活发生在新皮层上态转换之后[37]。这种时间上的关系可能会潜在地允许皮质皮质通过内嗅皮质起作用,从而影响海马体中哪些模式被重新激活。这样的输入可以提供部分触发,从而使CA3模式完成可以恢复最近的存储轨迹。反过来,在SWR发放过程中,这些完整的CA3模式可以通过CA1区域募集特定皮质组件的子集。确实,已经显示出,在某些从海马体直接接受输入的大脑区域中,单位在海马体SWR后持续发放——这些区域包括内嗅皮层[91],前额叶皮层[92]和腹侧纹状体[82]。
Conclusions
在许多皮层区域,即使不是全部,在随后的睡眠中神经元活动的清醒模式也会自发复现。确实,任何具有兴奋性循环性侧支系统的皮质回路都可能会存储清醒模式以重新激活。
海马体似乎特别准备协调整个大脑的重新激活:它处于关联处理的皮质层次结构的末端,可以访问高度精细的多模式表示。此外,海马体CA3中的锥体细胞集合紧密地相互连接,使其能够关联,存储和调用大量复杂的模式[23]。最后,它具有在联机和脱机模式之间快速切换的能力,这似乎分别适合于记忆形成和巩固。这与这样的想法是一致的,即最初由模态特异性新皮层回路处理的清醒经验可以被关联地绑定在海马体中,从而在记忆轨迹的不同组件之间建立了长期联系。正是这种组成成分的完整关联构成了一种回合记忆,看来这种关联的长期巩固需要海马体重新激活,以便协调下游皮层区域中相关组成成分的逐步募集。海马体中的SWR网络爆发模式以及与之相关的皮质事件可能反映了这种募集的机制,因此可以为复杂记忆轨迹的受控巩固提供底物。
尽管仍然存在许多问题(Box 2),但上面回顾的证据强烈表明,与SWR相关的海马体重新激活是一种强大的现象,具有实现回合记忆巩固的必要属性。光遗传学提供对海马体处理的更大的时间控制[93]应该为重新激活在巩固中的作用以及这种现象的潜在机制提供更多确定的证据。